Chromatin-Remodellierung
| RNA-Polymerasen |
Eukaryotisches Gen |
Regulation der Transkription |
Introns, Exons und Splicing
Hinweis: Im Atelier finden Sie die CD "The Nature of Genes". Mittels Tutorials und Aufgaben werden die wichtigsten Themen der Molekularbiologie leicht verständlich vermittelt.
Die Transkription bei Eukaryonten verläuft nach den gleichen Prinzipien wie die Transkription bei den Prokaryoten; es gibt jedoch einige wichtige Unterschiede:
Die DNA eukaryotischer Zellen ist als Chromatin organisiert. Die Transkription einzelner Gene bedingt vorgängige Veränderungen in der lokalen Chromatinstruktur, welche Gene für den Transkriptionsapparat zugänglich machen:
Im Gegensatz zu prokaryotischen Zellen besitzen eukaryotische Zellen nicht nur eine, sondern gleich drei verschiedene RNA-Polymerasen: RNA-Polymerase I, II und III.
Wie die prokaryotische RNA-Polymerase sind sie alle sehr gross und aus mehreren Untereinheiten aufgebaut. Und wie jede andere Polymerase synthetisieren sie neue Ketten in 5' -> 3' Richtung. Diese RNA-Polymerasen erkennen Promotoren, welche sich strukturell stark voneinander unterscheiden.
Wir wollen nur die RNA-Polymerase II-Gene näher betrachten.
Eukaryotisches protein-kodierendes Gen
Das Genom des Menschen enthält schätzungsweise 30'000-50'000 Gene, welche für Eiweisse kodieren. In ihrem grundsätzlichen Aufbau gleichen sie den prokaryotischen Genen:
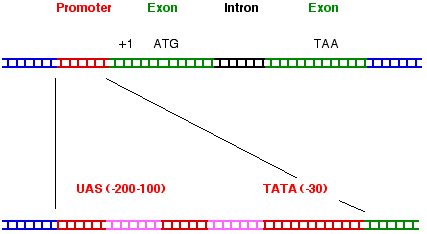
Die Promoter-Region: Wie die prokaryotischen Promotoren enthalten auch die RNA-Polymerase- II-Promotoren Bindungsstellen für die RNA-Polymerase und für Proteine, die diese Bindung modulieren können (UAS, upstream activating sequence). Die TATA-Box, analog der Pribnow-Box in Prokaryoten, bestimmt den Transkriptions-Start bei Position +1. Viele RNA-Polymerase II-Promotoren haben noch eine Enhancer (Verstärker)-Region. Sie kann, wie in der Abbildung dargestellt, vor dem eigentlichen Promoter liegen, kommt aber nicht selten hinter dem Gen oder gar mitten im Gen (z.B. in einem Intron) vor. Die Enhancer-Region stimuliert die Transkription des Gens mittels Proteinen, welche an diese DNA-Sequenz binden.
Transkribierte Region: Der Anfang der transkribierten Region, Position +1, heisst cap site, weil nach dem Beginn der Transkription die entstehende mRNA am ersten Nukleotid durch Anhängen der Cap-Struktur modifziert wird. Die Cap-Struktur wird durch Uebertragung von GTP auf die 5' terminale Base und Methylierung von Guanin in Position 7 synthetisiert. Die Cap-Struktur schützt die mRNA gegen Abbau durch Nukleasen und ist wichtig für die Translation.
Termination: Nach dem Passieren der Poly(A)-Addierungs-Sequenz AAUAAA auf der neusynthetisierten RNA pausiert die RNA-Polymerase II, assoziierte Proteine erkennen die Sequenz AAUAAA und die Transkription stoppt. Ein Enzymkomplex bindet an die Poly(A)-Addierungs-Sequenz AAUAAA der RNA, schneidet sie 10 - 15 Nukleotide weiter hinten (3' -Richtung) und hängt 50-150 Adenosine (Poly(A)-Sequenz) an.
Anordnung von Genen: In höheren Eukaryoten finden wir sehr oft Gene, die für das gleiche oder sehr ähnliche Proteine kodieren, sog. Gen-Familien. In der Regel werden nur wenige Gene, oft gar nur ein Gen aus einer Gen-Familie exprimiert. Die nicht-funktionellen Mitglieder der Gen-Familie werden als Pseudogene bezeichnet. Eukaryoten haben, im Gegensatz zu Prokaryoten, Gene nicht in Operons organisiert.
Genom-Grössen: Vergleicht man die Genom-Grössen von Zellen, so stellt man fest, dass sie mit zunehmendem Entwicklungsstand während der Evolution zunehmen. Eigenartigerweise stimmt diese Beziehung innerhalb der Eukaryoten nicht generell. So variiert die Genom-Grösse innerhalb der Amphibien von 109 bis 1011 Basenpaaren. Zudem sind die Genome grösser, als man aus der Anzahl Gene (10'000 - 100'000) in eukaryotischen Zellen erwarten würde. Aus dieser Beobachtung ergibt sich, dass nur ein kleiner Teil des Genoms von höheren Eukaryoten für Protein kodiert. Was ist die restliche DNA und welche Funktion hat sie? Die Genome von höheren Eukaryoten enthalten neben den einzelnen Genen noch hoch- und mittel-repetierte DNA-Sequenzen. Zu den hochrepetierten Sequenzen gehören DNA-Abschnitte von 300 - 1000 Basenpaaren Länge, die bis zu 100'000-fach wiederholt vorkommen. Ihre Funktion ist nicht bekannt. Zu den mittel-repetierten Sequenzen gehören die Gene für ribosomale RNA und tRNAs. Diese Gene sind repetiert, weil grosse Mengen der entsprechenden Gen-Produkte von der Zelle benötigt werden.

Die Regulation der Transkription ist aus mehreren Gründen von grosser Bedeutung:
|
1. |
Der gesamte Prozess der Transkription und Translation ist sehr energieaufwendig und sollte daher auf das notwendige Mass beschränkt sein. |
|
2. |
Durch verstärkte oder verminderte Synthese eines Enzyms kann dessen Konzentration in der Zelle erhöht resp. erniedrigt und dadurch der Stoffwechsel reguliert werden. |
|
3. |
Bei höheren Organismen sind viele Merkmale, die im Verlaufe der Entwicklung ausgebildet werden, genetisch determiniert. Die hierfür verantwortlichen Gene müssen also in bestimmten Entwicklungsphasen "angeschaltet" werden, um ihre Funktion zu erfüllen. |
Die Genregulation bei Eukaryoten wurde erst durch die Methoden der Analyse, Sequenzierung und Veränderung (Deletion, Mutation) definierter Genabschnitte einer gezielten Untersuchung zugänglich. Ein Ansatz bestand zunächst darin, DNA Strukturen in der Umgebung gleichartig regulierter Gene miteinander zu vergleichen und strukturell ähnliche Abschnitte zu benennen und ihre Funktion zu analysieren.
Durch Verwendung der geschilderten Methoden ist es gelungen, DNA-Abschnitte zu beschreiben, die die Expression eines Gens beeinflussen. Die TATA-Box und UAS als Teil der Promotorstrukturen wurde bereits genannt. Eine Sequenzklasse, die zunächst bei Eukaryonten-Viren beschrieben, dann aber auch für zelluläre Gene gefunden wurde, sind die Enhancer-Elemente, welche die Aktivität der Promotoren drastisch steigern.
Wichtige Tatbestände zur Genregulation:
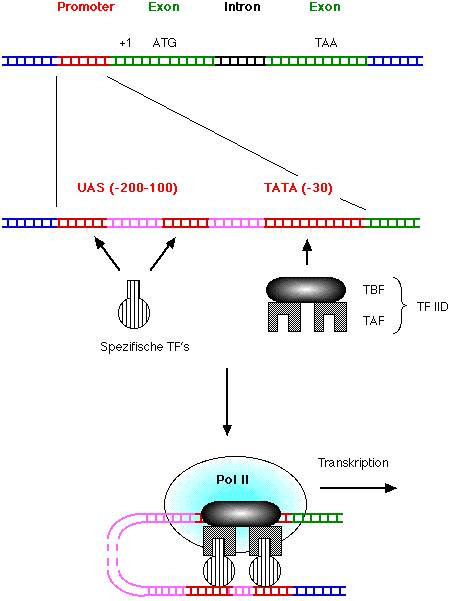
Fig.: TF, Transkriptionsfaktor; UAS, upstream activating sequence; TBF, TATA-binding factor; TAF, TATA-associated factor; Pol II, RNA-Polymerase II.
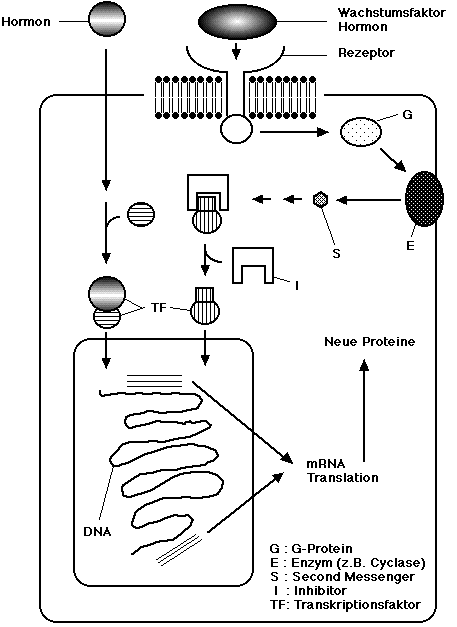
Figur: Die Bindung des Hormons an den Rezeptor führt zur Aktivierung einer Signaltransduktionskette.
Die Vorstufen der meisten eukaryotischen mRNA-Moleküle
enthalten die kodierende Sequenz nicht in ununterbrochener
Reihenfolge. Es sind vielmehr ein oder mehrere Stücke nicht in
Protein zu übersetzende Basensequenzen eingeschoben. Dies
bedeutet, dass die Gene, an denen diese Prä-mRNA synthetisiert
wurde, durch entsprechende DNA-Abschnitte unterbrochen sind. Man
bezeichnet die nicht-kodierenden Sequenzen als Introns und die
kodierenden Sequenzen als Exons.
Das primäre Transkript der Polymerase II ist eine hochmolekulare RNA, die auch Prä-mRNA genannt wird. Die Umwandlung (Prozessierung) der Prä-mRNA zur reifen mRNA beginnt damit, dass am 5'-Ende die sog. Cap-Struktur angehängt wird (siehe oben).
Die zunächst transkribierten Intronabschnitte werden im Verlaufe der Reifung der Prä-mRNA herausgeschnitten, und die kodierenden Abschnitte der mRNA werden zusammengefügt. Diesen Vorgang, den man in Analogie zum Spleissen von Tauwerk als das Splicing der RNA bezeichnet.
Wir unterscheiden 3 Typen (I,II und III) von Introns in eukaryotischen Genen. Sie unterscheiden sich vor allem in der Art und Weise, wie sie aus der Vorläufer-RNA entfernt werden. Nur Typ III kommt in protein-kodierenden Genen vor:
Eine interessante ungeklärte Frage ist diejenige nach der Entstehungsweise von Introns während der Evolution.
Es gibt Prä-mRNAs, welche nicht immer gleich gespleisst werden, d.h. es können von einer hnRNA mehrere unterschiedliche mRNAs angefertigt werden. Dieses Phänomen bezeichnen wir als "Alternatives Spleissen" (alternative splicing). Alternatives Spleissen wird reguliert und kann die kodierende Kapazität eukaryotischer Genome beträchtlich erhöhen.
Fertig modifizierte RNA wird als mRNA (messenger RNA) bezeichnet. Sie wird vom Kern ins Zytoplasma der Zelle transportiert, wo sie von Ribosomen gebunden und transliert wird.
Hinweis: Sie können sich im Atelier das Video "Transkription bei Eukaryonten" ansehen.
|
|
|
Transkription bei Prokaryonten |